ukončenie je konečná fáza replikácie DNA. Predchádza mu začatie a predĺženie. Včasné ukončenie replikácie môže mať za následok expresiu skrátených proteínov a teda mutáciu.
Čo je ukončenie?

Počas replikácie alebo reduplikácie sa DNA genetického nosiča informácií rozmnožuje v jednotlivých bunkách. K duplikácii dochádza podľa polokonzervatívneho princípu a zvyčajne vedie k presnej duplikácii genetickej informácie. Replikácia sa spúšťa počas fázy syntézy, pred fázou mitózy, a teda prebieha pred delením bunkového jadra.
Na začiatku replikácie sa dvojreťazec DNA rozdelí na jednotlivé vlákna, na ktorých sa vytvoria nové komplementárne vlákna. Každý reťazec DNA je určený sekvenciou báz opačného reťazca. Replikácia DNA prebieha v niekoľkých fázach. Ukončenie je tretia, a teda posledná fáza replikácie. Pred ukončením predchádza začatie a predĺženie.
Synonymom pre výraz ukončenie v tejto súvislosti je označenie Ukončenie fázy, Ukončenie tu znamená „prerušenie“ alebo „ukončenie“. Počas ukončenia sa novoformované vlákno mRNA oddelí od skutočnej DNA. Práca DNA polymerázy sa pomaly končí. Ukončenie replikácie DNA by sa nemalo zamieňať s ukončením replikácie RNA.
Funkcia a úloha
Vo fáze replikácie iniciácie sa uskutočňuje predovšetkým regulácia replikácie. Stanoví sa počiatočný bod replikácie a dôjde k tzv. Aktivácii. Po iniciácii sa začne polymerizácia, počas ktorej prebieha predlžovacia fáza. Enzým DNA polymeráza separuje komplementárne vlákna DNA na jednotlivé vlákna a číta bázy jednotlivých vlákien jeden po druhom. V tejto fáze dochádza k dvojdiskontinuálnemu zdvojeniu, ktoré zahŕňa ďalšiu fázu aktivácie.
Až po začatí a predĺžení nasleduje fáza ukončenia v rámci replikácie. Ukončenie sa líši od formy života k forme života. V eukaryotoch, ako sú ľudia, je DNA štruktúrovaná v kruhu. Zahŕňa tiež terminačné sekvencie, ktoré zodpovedajú dvom rôznym sekvenciám, z ktorých každá je relevantná pre replikačnú vidlicu.
Ukončenie zvyčajne nespúšťajú špeciálne mechanizmy. Hneď ako spolu prebehnú dve replikačné vidlice alebo keď DNA skončí, replikácia sa v tomto bode automaticky ukončí. Replikácia sa ukončí automatickým mechanizmom.
Terminačné sekvencie sú kontrolné prvky. Zabezpečujú, aby sa replikačná fáza dostala do špecifického koncového bodu kontrolovaným spôsobom napriek rozdielnym rýchlostiam replikácie v dvoch replikačných vidliciach. Všetky terminačné miesta zodpovedajú väzbovým miestam pre proteín Tus, „látka využívajúca terminus“. Tento proteín blokuje replikatívnu helikázu DnaB a tak zastavuje replikáciu.
V eukaryotoch zostávajú replikované kruhové vlákna navzájom spojené aj po replikácii. Spojenie zodpovedá koncovému bodu. Až po delení buniek sú oddelené rôznymi procesmi, a preto ich možno rozdeliť. Zdá sa, že spojenie, ktoré zostáva až po delení buniek, hrá úlohu v riadenej distribúcii.
Pri konečnej separácii kruhov DNA hrajú úlohu dva hlavné mechanizmy. Pri separácii sa zúčastňujú enzýmy, ako je napríklad topoizomeráza typu I a typu II. Nakoniec pomocný proteín rozpoznáva stop kodón počas terminácie. To spôsobuje, že polypeptid spadne z ribozómu, pretože nie je k dispozícii žiadna t-RNA s vhodným antikodónom pre stop kodón. Nakoniec sa ribozóm rozkladá na svoje dve podjednotky.
Choroby a choroby
Všetky procesy na duplikáciu genetického materiálu v zmysle replikácie sú komplikované a vyžadujú vysoké množstvo látok a energie v bunke. Z tohto dôvodu sa môžu ľahko vyskytnúť spontánne chyby replikácie. Ak sa genetický materiál zmení spontánne alebo je vyvolaný zvonku, hovoríme o mutáciách.
Chyby replikácie môžu viesť k chýbajúcim základniam, môžu byť spojené so zmenenými základňami alebo môžu byť spôsobené nesprávnym párovaním báz. Okrem toho delécia a inzercia jedného alebo viacerých nukleotidov do dvoch reťazcov DNA môže viesť k chybám replikácie. To isté platí pre pyrimidínové diméry, zlomy vlákien a chyby zosieťovania v reťazcoch DNA.
V prípade chyby replikácie sú k dispozícii samostatné opravné mechanizmy. Mnohé z uvedených chýb sú v najväčšej možnej miere korigované DNA polymerázou. Presnosť replikácie je pomerne vysoká. Miera chybovosti je iba jedna chyba na nukleotid, ktorú možno pripísať rôznym kontrolným systémom.
Napríklad kontrolný mechanizmus eukaryotických buniek je známy ako nezmyslom sprostredkovaný rozpad mRNA, ktorý dokáže rozpoznať nežiaduce stop kodóny v mRNA a tak zabrániť skráteným proteínom v nájdení expresie.
Predčasné stop kodóny v mRNA sú spôsobené génovými mutáciami. Takzvané nezmyslové mutácie alebo alternatívne a chybné zostrihy môžu mať za následok skrátenie proteínov, ktoré sú ovplyvnené funkčnými stratami. Kontrolné mechanizmy nemôžu chyby vždy opraviť.
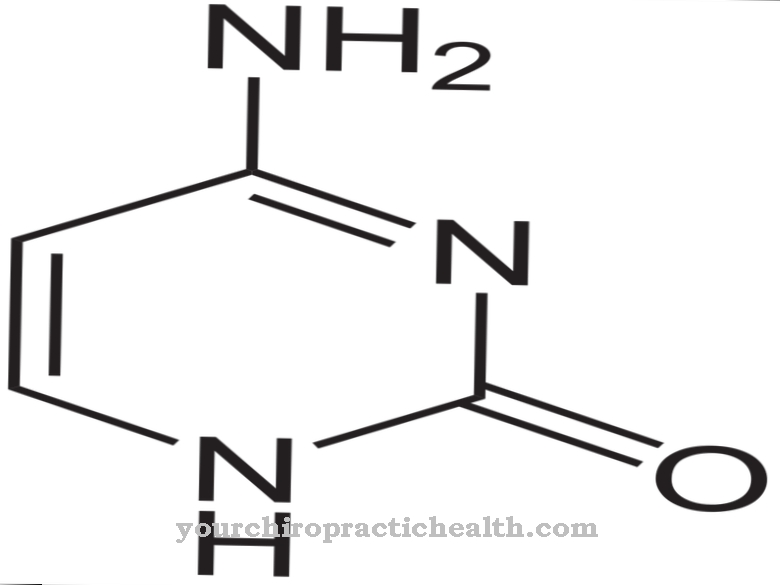
Autozomálna recesívna dedičná porucha β-talasémia má tri rôzne formy: prvou je homozygotná talasémia, závažné ochorenie, ktoré sa dá vysledovať až k vašej nezmyselnej mutácii. Heterozygotná talasémia je miernejším ochorením, pri ktorom sa nezmyslové mutácie nachádzajú iba v jednej kópii génu ß-globínu. Prostredníctvom mechanizmu nezmyslom sprostredkovaného rozpadu mRNA môže byť mRNA chybného génu degradovaná do tej miery, že sú exprimované iba zdravé gény.
Pri heterozygotnej talasémii a teda miernej forme ochorenia je nezmyslová mutácia v poslednom mRNA exóne, takže kontrolné mechanizmy nie sú aktivované. Z tohto dôvodu sa popri zdravom ß-globíne tvorí aj skrátený ß-globín.Erytrocyty s defektnou ß-globínom hynú.
Iným príkladom zlyhania kontrolného mechanizmu je Duchennova svalová dystrofia, ktorá je tiež spôsobená nezmyslovou mutáciou v mRNA. V tomto prípade kontrolný mechanizmus štiepi mRNA, ale tak spôsobuje úplnú stratu takzvaného dystrofínového proteínu.





.jpg)
.jpg)


