epitelový-mezenchymálny prechod, tiež EMT "transformácia epitelových buniek do mezenchymálnych buniek". Táto transformácia má veľký význam pre embryonálny vývoj. Tento proces však tiež zohráva kľúčovú úlohu pri vývoji metastáz v karcinómoch.
Čo je to epitelový-mezenchymálny prechod
Epitelový-mezenchymálny prechod je premena už diferencovaných epitelových buniek na nediferencované mezenchymálne kmeňové bunky. Tento proces má osobitný význam počas embryonálneho vývoja.
V rámci tejto transformácie sa epitelové bunky uvoľňujú zo svojej väzby a môžu v tele migrovať. Pritom prechádzajú bazálnou membránou. Suterénna membrána oddeľuje epitel, gliové bunky a endotel z bunkového priestoru podobného spojivovým tkanivám. Ako nediferencované multipotentné kmeňové bunky migrované bunky zasahujú do všetkých oblastí vyvíjajúceho sa organizmu a môžu sa opäť diferencovať na akýkoľvek typ bunky.
Epitelové bunky tvoria tzv. Epitel, čo je spoločný názov pre žľazové a krycie tkanivo. Mezenchým obsahuje želatínové a embryonálne spojivové tkanivo, z ktorého sa vyvíjajú kosti, chrupavka, hladké svaly, srdcové svaly, obličky, kôra nadobličiek, krvotvorný systém s krvou a lymfatickými cievami a retikulárne, pevné a voľné spojovacie tkanivo.
Funkcia a úloha
Epitelový-mezenchymálny prechod je dôležitým procesom počas embryogenézy. Počas tejto doby dochádza k zvýšenému rastu, na ktorom sa podieľajú všetky bunky tela. Epitelové bunky, ktoré už boli diferencované, sú tiež zahrnuté v týchto rastových procesoch. Aby to však mohli urobiť, musia byť prevedené späť na multipotentné kmeňové bunky.
Najintenzívnejší rast nastáva počas prvých ôsmich týždňov tehotenstva. Skutočný proces embryogenézy začína okolo šiesteho dňa tehotenstva po tzv. Klíčivom štádiu (vývoj buniek) a trvá až do konca ôsmeho týždňa tehotenstva. V tejto fáze má veľký význam prechod epitelu a mezenchýmu, pretože všetky orgány sa teraz vytvárajú. Mnoho epitelových buniek tu opäť stráca svoju diferenciáciu a prichytenie. Migrujú cez suterénnu membránu a sú distribuované v tele. Tam sa správajú znova ako normálne multipotentné kmeňové bunky a podliehajú obnovenej diferenciácii na rôzne typy buniek.
Samozrejme sa môžu opäť diferencovať na epitelové bunky. Aby sa to dosiahlo, je potrebné najprv zredukovať bunkové kontakty a zrušiť polaritu epitelových buniek. Bunkovým kontaktom sa rozumie kohézia buniek pomocou takzvaných adhéznych molekúl. E-kadherín je dôležitou adhéznou molekulou. E-kadherín je transmembránový glykoproteín, ktorý je závislý od iónov vápnika. Spája medzi sebou epitelové bunky a zaisťuje polaritu buniek a prenos signálu. Počas embryogenézy je aktivita E-kadherínu znížená. To vedie k uvoľneniu bunkovej štruktúry. Zároveň zmizne aj polarita buniek.
Epitelové bunky majú takzvanú apikálnu (vonkajšiu) aj bazálnu stranu smerujúcu k podkladovému tkanivu. Vonkajšia strana je na povrchu kože a slizníc, zatiaľ čo bazálna strana je spojená s väzivovým tkanivom umiestneným pod bazálnou laminou. Obe strany majú odlišné funkčné a štrukturálne rozdiely, a tak zabezpečujú morfológiu orgánov. Embryogenéza si však vyžaduje rýchle zmeny a flexibilitu buniek, aby sa dokázala rýchlo prispôsobiť rastovým procesom.
Po ukončení embryogenézy stráca epiteliálno-mezenchymálny prechod pre organizmus svoj význam.
Choroby a choroby
Prechod epitelu a mezenchýmu (EMT) je pre organizmus prospešný iba počas veľmi krátkeho obdobia embryogenézy. Po búrlivej rastovej fáze sa bunky diferencujú. Potom už nie je potrebné veľké množstvo multipotentných kmeňových buniek. Preto je tento proces zakázaný.
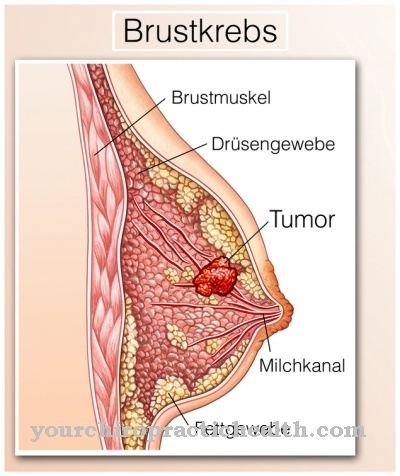
Ak je epitelový mezenchymálny prechod aktivovaný po ukončení embryogenézy, zvyčajne sa to stáva v súvislosti s malígnymi nádorovými ochoreniami. EMT je zodpovedný za vývoj metastáz v kontexte rakoviny. Tento proces je podobný procesu embryogenézy. Celkovo je to komplexný proces založený na genetických regulačných mechanizmoch, ktoré ešte nie sú úplne objasnené. Mnoho zodpovedných génov je aktívnych iba počas embryonálneho vývoja. Potom sú vypnuté. Možnou príčinou obnovenej aktivácie týchto génov by mohla byť zvýšená regulácia transkripčného faktora Sox4. Zodpovedajúce výsledky výskumu boli prezentované na Bazilejskej univerzite. Sox4 zase aktivuje množstvo ďalších génov, ktoré sa podieľajú na epitelovo-mezenchymálnom prechode.
Nečinnosť zodpovedajúcich génov je údajne založená na ich nečitateľnosti v dôsledku poťahovania určitými proteínmi (histónmi). Gén Sox4 je však zodpovedný za tvorbu enzýmu nazývaného Ezh2. Je to metyltransferáza, ktorá spôsobuje metyláciu zodpovedajúcich histónov. Ostatné zapojené gény sa stanú opäť čitateľnými a aktivujú epitelový mezenchymálny prechod.
Zmena v genetickom materiáli nastáva v rakovinovom nádore, a tak poskytuje príčinu úplného dediferencovania rakovinových buniek. Bez epitelového-mezenchymálneho prechodu by rakovina rástla iba v mieste pôvodu a nerozširovala sa. Tvorba metastáz však robí nádor zvlášť malígny a agresívny. Preto pracujeme na vývoji liekov, ktoré inhibujú tvorbu metyltransferázy Ezh2. Zodpovedajúce lieky už boli vyvinuté, stále sa však testujú. Obmedzenie tvorby metastáz by na jednej strane znížilo agresivitu rastu rakoviny a na druhej strane by otvorilo príležitosť liečiť predtým beznádejné prípady.





.jpg)









.jpg)